Zielsetzung
Mit Hilfe von Extinktionsmessungen im Photometer und high pressure liquid chromatography (HPLC) sollen die einzelnen Photosynthesepigmente in ihrem Absorptions- und Fluoreszenzverhalten und quantitativ charakterisiert werden
Versuchsdurchführung
Aus einer Mutanten Tabakpflanze wurde ca. 20g Blattmaterial mit ca. 200ml Isolationsmedium (50mM Tricin; pH: 7,8; 330mM Sorbitol) im Mixer homogenisiert. Das Homogenat wurde filtriert und mehrmals zentrifugiert und das Pellet jeweils in einem Schockmedium ohne Sorbitol resuspendiert. Als Ergebnis erhält man eine Thylakoidsuspension.
Zur Herstellung einer reinen Pigmentsuspension wurden 400\(\mu\)l Thylakoidsuspension mit 1,6ml basischem Aceton (10\(\mu\) M KOH) versetzt und zentrifugiert. Die organische Phase enthält die gewünschte Pigmentsuspension.
Phaeophytinbildung
100 \( \mu\)l der Pigmentlösung wurden mit 200 \( \mu\)l Methanol verdünnt und anschließende tropfenweise mit 1 M HCl vesetzt.
Ergebnis und Auswertung
Nach der Zugabe von HCl verfärbte sich die Pigmentlösung von grün nach braun. Phaeophytin ist ein Chlorophyllmolekül, bei dem das zentrale \(Mg^{2+} \) durch \(2H^+ \) ersetzt wurde. Pheophytin besitzt durch die 2 Protonen ein anderes Absorptionsspektrum, wodurch eher rotes Licht transmittiert wird. Durch die Suspension mit Chlorophyll und anderen Pigmenten ergibt sich eine bräunliche Färbung. Die biologische Bedeutung von Phaeophytin besteht in der Elektronentransportkette als Elektronencarrier.
Gesamtproteingehalt mittels Bradford-Reagenz
Zur Bestimmung des Gesamtproteingehaltes wird jeweils 10 \(\mu\)l verschiedener Verdünnungsstufen der Thylakoidsuspension mit 1 ml Bradford-Reagenz vermischt und bei 595 nm im Photometer die Extinktion bestimmt. Als Bezugsgröße diente der Blindwert. Zur Berechnung der jeweiligen Proteingesamtmenge wurde die Eichgerade mit BSA-Lösung aus dem Skript verwendet. Die Proteinmenge wurde dann mit der angegebenen Verdünnung auf die Ausgangslösung hochgerechnet. \[ E_{\lambda{}959}=0,5593*BSA [mg/ml] \]
| Thylakoide | Verdünnung | \(E_{\lambda{}595}\) | Proteine |
|---|---|---|---|
| \(0 \mu l\) | 0:1 | 0 | 0 |
| \( 10 \mu l\) | 1:100 | 0,055 | 9,8 |
| \( 50\mu l\) | 1:20 | 0,296 | 10,6 |
| \( 100 \mu l\) | 1:10 | 0,673 | 12,0 |
Mittelwert: \[ \bar{X}=\mathbf{10,8 \frac{g}{l}} \]
In der Thylakoidmembran sind die Pigmente an transmembranen Pigmentproteinkomplexen gebunden. Dazu gehören die Proteine der Photosysteme I und II mit den integrierten Lichtsammelkomplexen LHC. Ihre Funktion besteht im größten Teil darin, die einzelnen Pigmente und Co-Faktoren wie z.B. Plastoquinon oder Mn-Cluster zu positionieren und so den Transfer der Excitonenenergie zu ermöglichen. Weitere Proteine aus dem Elektronentransfer (Cytochrom \(b_6\)/f-Komplex) und der ATP-Synthese (\(F_0/F_1 \) ATPase) sind ebenfalls in de Thylakoidmembran lokalisiert.
Abweichungen in der Proteinkonzentration können durch Verunreinigungen der Pigmentlösung entstehen. So können Reste von Stärke die Extinktion erhöhen. Außerdem ist die Bradfordreaktion eine Gleichgewichtsreaktion und die stabile Komplexbildung des Farbstoffes mit den hydrophoben Seitenketten der Proteine benötigt eine gewisse Zeit. Außerdem wurde keine eigene Eichgerade angefertigt, wodurch die verwendete BSA-Gerade aus dem Skript für die Probe nicht vollkommen repräsentativ sein könnte.
Quantitative spektrophotometrische Bestimmung
Die hergestellte Pigmentsuspension enthält Chlorophyll-a und Chlorophyll-b und eine Reihe von verschiedenen Carotinoiden. Zur Bestimmung der Pigmentkonzentrationen wird die Extinktion bei verschiedenen Wellenlängen gemessen. Aus dem Beer-Lamber-Gesetz lässt sich für die gemessene Extinktion folgender Zusammenhang aufstellen: \[ E_{\lambda 1}= [A] * \epsilon_{A_{\lambda 1}}d + [B] * \epsilon_{B_{\lambda 1}}d \]
Dabei sind die Extinktionskoeffizienten der einzelnen Substanzen bekannt und durch die Messung bei mehreren Wellenlängen lässt sich ein Gleichungssystem für die gesuchte Konzentration aufstellen: \[ [Chl a] = 12,25*E_{663,6} - 2,55*E_{646,6}[\mu g/ml] \] \[ [Chl b] = 20,31*E_{646,6} - 4,91*E_{663,6}[\mu g/ml] \] \[ [Chl a+b] = 17,76*E_{646,6} + 7,34*E_{663,6}[\mu g/ml] \] \[ [Car] = \frac{1}{198}* \big( 1000*E_{470} - 1,91*[Chl a] - 95,15*[Chl b] \big)[\mu g/ml] \]
Die berechnete Pigmentkonzentration wird mit dem Verdünnungsfaktor 1:100 korrigiert und anschließend auf die vorher berechnete Proteinmenge bezogen.
Ergebnis und Auswertung
| Extinktion | Pigmente pro Protein | ||||||
|---|---|---|---|---|---|---|---|
| \(E_{470}\) | \(E_{646}\) | \(E_{663}\) | \(E_{720}\) | [Chl a] | [Chl b] | [Car] | |
| 1,538 | 0,620 | 1,547 | 0,010 | 0,161 | 0,046 | 0,048 | |
Qualitative und Quantitative Charakterisierung mittels HPLC
Für die HPLC wurden 200\(\mu\)l Pigmentlösung zentrifugiert und der Überstand für die HPLC-Analyse überführt.
Das Prinzip der Pigmenttrennung mittles high pressure liquid chromatography beruht auf dem unterschiedlichen Verhalten der mobilen und stationären Phase der einzelnen Substanzen auf Grund ihrer Polarität. Bei der reversed phase HPLC werden im Gegensatz zur Dünnschichtchromatografie Trennmaterialien eingesetzt, bei denen die mobile Phase weniger polar ist als die stationäre. Aus diesem Grund werden in der RP HPLC die unpolaren Substanzen besonders gut zurückgehalten.
Das Auftrennverhalten der einzelnen Substanzen ist deshalb entgegengesetzt zur DC. Aus dem Elutionsprofil lässt sich direkt die Polarität der einzelnen Pigmente schlussfolgern. Am unpolarsten ist das \( \beta \)-Carotinoid als reiner Kohlenwasserstoff und läuft deshalb auch als letztes durch die Trennsäule, vorausgegangen von den Chlorophyllen, wobei Chl a unpolarer ist als Chl b mit der zusätzlichen Carbonylgruppe. Davor kommen die sauerstoffhaltigen Xanthophylle. Das polarste Pigment ist schließlich das Neoxanthin mit drei Hydroxygruppen und einer Epoxygruppe, welches als erstes aus der Säule gewaschen wird.
Ergebnisse und Auswertung
Aus der integrierten Peakfläche multipliziert mit einem Faktor für bekannte Standardkonzentrationen ergibt sich die Konzentration der Pigmente im Einspritzvolumen. Unter Berücksichtigung der Verdünnung von 1:5 lassen sich die Angaben auf die Orginalprobe und die Proteinmenge zurückrechnen. Bsp: \[ Neo = 3856790 *82*10^{-6}* \frac{1}{25}*5*\frac{10^{-9}}{10^{-6}}*10,8^{-1} = 5,85*10^{-3} \]
Die Angaben sind dabei in Gramm Pigment pro Gramm Protein.
| Pigment | Zeit [min] | Fläche | Konzentration [ng/25\(\mu\)l] | pro Protein |
|---|---|---|---|---|
| Neoxanthin | 8,76 | 3856790 | 1581,30 | \(5,85*10^{-3}\) |
| Violaxanthin | 12,25 | 3682464 | 1141,55 | \(4,25*10^{-3}\) |
| Antheraxanthin | 15,41 | 303417 | 95,60 | \(0,35*10^{-3}\) |
| Lutein | 17,51 | 13569384 | 4070,8 | \(15,10*10^{-3}\) |
| Zeaxanthin | 18,26 | 639829 | 230,35 | \(0,85*10^{-3}\) |
| Chl-b | 22,43 | 27844772 | 13922,40 | \(51,55*10^{-3}\) |
| Chl-a | 24,67 | 48000694 | 34320,50 | \(127,1*10^{-3}\) |
| \(\beta\)-Carotin | 31,49 | 8300259 | 2324,05 | \(8,60*10^{-3}\) |
Das Verhältnis von Chl-b zu Chl-a beträgt: 1:2,47 Das molare Verhältnis der Gesamtcarotinoide zum Gesamtchlorophyll berechnet sich nach Formel(3) und beträgt 3,1:1. Der Anteil der einzelnen Xanthophylle beträgt:
- Vio:Xanthophylle: 0,78
- Ant:Xanthophylle: 0,06
- Zea:Xanthophylle: 0,16
Xanthophylle sind chemisch gesehen Carotinoide mit sauerstoffhaltigen Gruppen. In den Pflanzen stellt Zeaxanthin ein wichtiges Schutzpigment vor Photooxidation dar. Im sogenannten Zeaxanthinzyklus wird durch Lichtenergie und NADPH-Verbrauch Violaxanthin über Antheraxanthin zu Zeaxanthin unter Bildung von Dehydroascorbat. Dabei wird wahrscheinlich Extinktionsernergie des Chlorophylls in Wärmeenergie umgewandelt und somit das Chl vor Photooxidation durch zu hohe Lichtintensität geschützt. Der hohe Anteil von Violaxanthin und der nur sehr geringe Zeaxanthin-Anteil lassen sich so interpretieren, dass die Pflanze in der Zeit vor der Experiment keinen hohen Lichtintensitäten ausgesetzt war und deshalb den Zeaxanthin-Zyklus nicht im hohen Maße durchlaufen hat.
Das Chlorophyllverhältnis von Chl-a zu Chl-b beträgt im HPLC-Verfahren 1:2,47 was beim Vergleich mit Literaturangaben dem Verhältnis von Chloroplasten unter Starklicht entspricht. Bei einer ausreichenden Beleuchtung bilden sich Chloroplasten mit reduzierten Thylakoiden und einer geringeren Stapeldichte. Im Gegensatz dazu ist bei Starklicht der Gehalt an Carotinoiden zum Schutz vor Photooxidation erhöht. Eine starke Beleuchtung der Pflanze lässt sich auch auf Grund des hohen Stärkegehaltes im Extrakt vermuten.
Der Vergleich der spektrometrischen Bestimmung mit der HPLC zeigt dagegen größere Abweichungen bei den Carotinoiden. Der Chlorophyllgehalt pro Protein unterscheidet sich bei beiden Methoden nur leicht und kann gut möglich in der Schwankung und im Fehlerbereich der Methode liegen. Das Verhältnis von Chl-a zu Chl-b liegt bei beiden Methoden im Bereich der Erwartung und für eine Signifikanz müssten weitere Messungen durchgeführt werden. Der Gehalt an Carotinoiden ist bei der HPLC-Messung jedoch viel größer als bei der spektroskopischen Messung. So stimmt das berechnete Verhältnis Chl:Car nicht mal in der Tendenz. Eine Möglichkeit das die Formel(2) auf Extinktionen bei \( E_\lambda = 663,3 nm \) und \( 646,6 nm \) beruht und wir bei 663nm und 646nm gemessen haben. Im Gesamten würden wir die HPLC-Methode als genauer erachten.
Fluoreszenz extrahierter Pigmente
Es wurden 100\(\mu\)l der Pigmentlösung unter UV-Licht zum Fluoreszieren gebracht und anschließend die Veränderung der Fluoreszenz nach Zugabe von Aqua dest. beobachtet.
Da die Chlorophylle in der Pigmentlösung nicht mehr durch Proteine und Thylakoide positioniert werden, können die Chlorophylle die absorbierte Energie aus dem \(S_1 \) Zustand nicht für die Photochemie an andere Chlorophylle weiterleiten. Die isolierten Chlorophylle in der Suspension geben deshalb die Absorbierte Energie vor allem durch Fluoreszenz wieder ab. Der \(S_2\)-Zustand ist sehr instabil, so dass nach einer Relaxation \(S_2 \rightarrow S_1 \) die Energie im stabilen \(S_1\)-Zustand durch Fluoreszenz abgegeben wird. Die Energiedifferenz von \(S_1 \rightarrow S_0 \) entspricht dabei einem Photon mit einer Wellenlänge im roten Farbbereich, wodurch Chlorophyll rot fluoresziert. Durch den Stokes-Shift ist das fluoreszierte Licht langwelliger als bei einer Absorption für eine \( S_0 \rightarrow S_1 \) Anregung. Durch Zugabe von Aqua dest. erlöscht die Fluoreszenz. Das ist dadurch begründet, dass Chlorophyll stark hydrophob ist und sich deshalb durch den hydrophoben Effekt zu Micellen zusammenlagert, um die Entropie der freien Wassermoleküle zu erhöhen. Dadurch treten die Chlorophylle wieder in engen Kontakt, wodurch sie ihre Energie wieder durch Resonanztransfer strahlungsfrei auf benachbarte Chlorophylle übertragen können.
Fehlerbetrachtung
Fehler können zum einen bei der Herstellung der Pigmentsuspensionen entstehen. So befand sich im Rohextrakt eine sehr hohe Konzentration an Stärke, welche vor allem die Messung im Photometer beeinflusst haben kann. Eine Verunreinigung oder ein Fehler bei der Verdünnung hätte sich somit in alle Versuchsteile fortgepflanzt. So kann es vor allem bei der Verdünnung mit Aceton zu Fehlern beim pipetieren gekommen sein, weil Aceton schnell aus der Pipettenspitze läuft. Des weiteren wurde keine eigene BSA-Standardkurve für den Bradfordtest erstellt, so dass es Fehler bei der Berechnung der Proteinkonzentration gekommen sein kann. Außerdem handelte es sich um eine Mutante-Tabakpflanze, so dass ein Vergleich mit Literaturwerten vielleicht nicht angebracht ist.
Absorptionsspektroskopie
Das Absorptionsspektrum der hergestellten Pigmentlösung und der (1:100 verdünnten) Thylakoidlösung wird im Wellenlängenbereich 400 nm bis 730 nm bestimmt. Die gewonnenen Messdaten wurden nun durch subtrahieren des Minimalwertes, und durch dividieren durch den langwelligen Maximalwert der Exktinktion normiert.
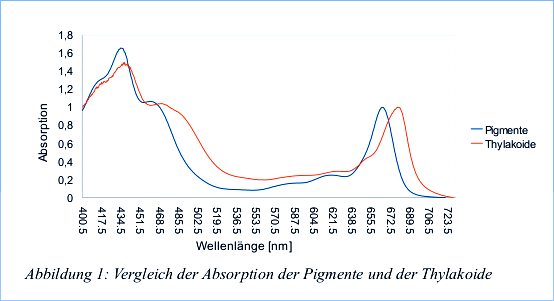
Auswertung
In der Pigmentlösung sind bei etwa 430 nm und bei etwa 460 nm deutlich die kurzwelligen Maxima (im blauen Spektralbereich) der Chlorophylle a und b zu erkennen. Diese Maxima der Chlorophylle gehen in die Absorptionsspektren der Carotinoide über, die im langwelligeren gelbgrünen Bereich absobieren. Diese zeigen allerdings keine deutlichen Maxima. Im langwelligen (roten) Bereich ist das Absorptionsmaximum von Chlorophyll a (bei etwa 660nm) wieder sehr deutlich zu sehen. Das Maximum von Chlorophyll b dagegen ist nicht sichtbar, da es vom Chlorophyll a überlagert wird. Merkwürdig sind das Maximum bei etwa 620 und seine Schulter, weil dort eigentlich weder Chlorophylle noch Carotinoide absorbieren sollten. Der Vorteil der unterschiedlichen Absorptionsmaxima ist , dass mehr Energie aufgenommen werden kann, da ein breiterer Spektralbereich abgedeckt wird. Die Funktion der Chlorophylle und Carotinoide ist unterschiedlicher Natur: Während Chlorophyll a auch im Reaktionszentrum des Photosystems vorkommt und dort die Photochemie betreiben, kommen Chlorophyll b und die Carotinoide nur in den LHC vor (allerdings auch dort Chlorophyll a), wo sie eine Licht sammelnde Funktion wahrnehmen. Die von den LHC aufgenommene Energie wird zum Reaktionszentrum weitergeleitet, um die Photosynthese anzutreiben. Zusätzlich zu ihrer Lichtsammelnden Funktion besitzen die Carotinoide noch eine Schutzfunktion. Wenn die Pflanze zu starken Lichtverhältnissen ausgesetzt ist, kann überschüssige Energie unter Wärmeabgabe von den Chlorophyllen an die Carotinoide weitergeleitet werden. Im Vergleich mit der Absorption der Thylakoidsuspension fällt auf, dass die Absorptionsbereiche viel breiter sind und das die Carotinoide deutlich mehr zu aborbieren scheinen. Im langwelligen Bereich ist auserdem zu sehen, dass das merkwürdige Maximum bei ca. 620 nm verschwindet und das Absorptionsmaximum von Chlorophyll a scheinbar ins noch langwelligere bis zu ca 680 nm verschoben wird. Diese Unterschiede sind dadurch zu erklären, dass in den intakten Thylakoiden die Photosynthesepigmente vernetzt sind, um eine effiziente Energieweiterleitung zu ermöglichen. Die höhere Absorpion in der Grünlücke ist damit zu erklären. Die Carotinoide können ihre Energie besser weiterleiten, und so auch mehr absobieren. Außerdem verbinden sich in den Thylakoiden die Photosynthesepimente mit Proeinen wodurch sich zum Beispiel das Absorptionsverhalten von zum Beispiel Chlorophyll a verschiebt. Dies ist besonders gut im roten Spektralbereich zu erkennen.
Fluoreszenzspektroskopie
In diesem Versuch werden die Fluoreszenz-Emissionsspektren der Thylakoide und der freien Pigemente bei anregendem Licht von 440 nm und 460 nm gemessen. Außerdem wurde das Fluoreszenzanregungsspektrum beider Suspensionen bei einer Emissionswellenlänge von 700 nm aufgezeichnet. Bei einem Emissionsspektrum wird bei einer definierten eingestrahlten Wellenlänge die Wellenlänge und Intensität des emmitierten Lichtes gemessen. Bei einem Anregungsspektrum dagegen wird die Wellenlänge des Anregenden Lichtes kontinuierlich verändert, und die Fluoreszenzintensität bei einer bestimmten Wellenlänge gemessen
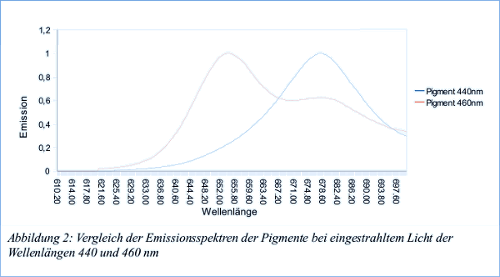
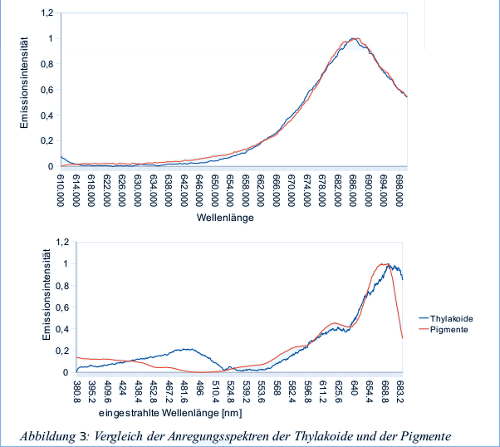
Während bei bei 440 nm Chlorophyll a angeregt wird regt Licht von 460 nm Wellenlänge Chlorophyll b an. Bei den reinen Pigmenten sind deshalb deutliche Unterschiede zwischen den Emissionsspektren zu beobachten, da bei 440 nm vor allem Chlorophyll a emitiert, bei 460 nm aber Chlorophyll b (Allerdings auch Chlorophyll a, wie in der Grafik zu sehen, was daran liegt, dass sich die Absorptionsbereiche der Chlorophylle überschneiden). Beim intakten Photosystem der Thylakoide hingegen wird die von Chlorphyll b aufgenommene Energie an Chlorophyll a weitergeleitet, weshalb die normierten Emissionsspektren hier genau gleich aussehen. Zum Anregungsspektrum der Pigmentlösung tragen Scheinbar alle Pigmente bei, Chlorophylle wie auch Carotinoide. Allerdings ist eine deutliche Blauverschiebung bei der Pigmentsupension gegenüber der Thylakoidsuspension zu sehen. Diese Blauverschiebung ist wie schon zuvor durch Komplexbildung der Pigmente mit Proteinen zu erklären, welche im intakten Photosystem (in den Thylakoiden) dafür sorgen, dass die Pigmente im etwas langwelligeren Bereich absorbieren.